 |
Kaji, Anker, Wirkner & Palmer, 2018.
|
Highlights
• The evolutionary history of remarkable snapping claws in shrimp is reconstructed
• Two novel claw-joint types—slip joints and torque-reversal joints—preceded snapping
• The transition “slip joint → torque-reversal joint → snapping” occurred in two families
• Subtle changes in joint form yielded dramatic changes in claw function (e.g., speed)
Summary
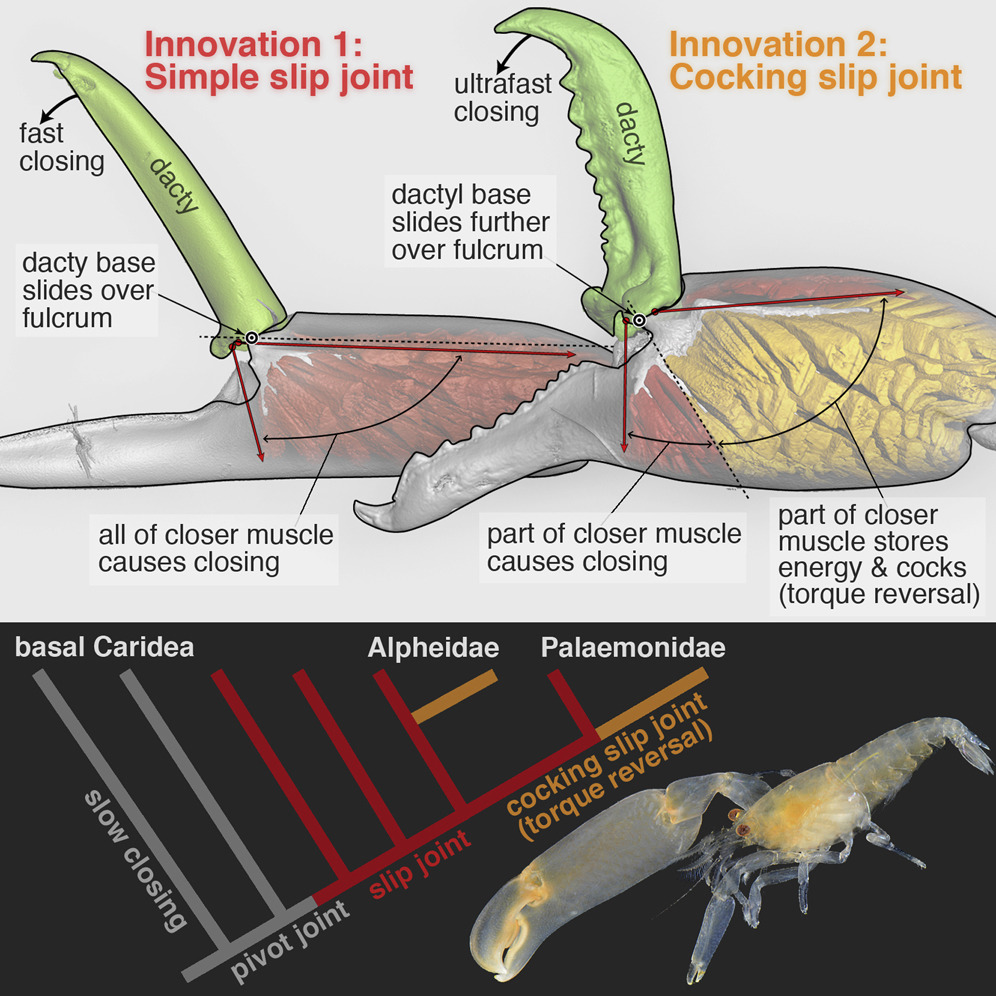
How do stunning functional innovations evolve from unspecialized progenitors? This puzzle is particularly acute for ultrafast movements of appendages in arthropods as diverse as shrimps, stomatopods, insects, and spiders. For example, the spectacular snapping claws of alpheid shrimps close so fast (∼0.5 ms) that jetted water creates a cavitation bubble and an immensely powerful snap upon bubble collapse. Such extreme movements depend on (1) an energy-storage mechanism (e.g., some kind of spring) and (2) a latching mechanism to release stored energy quickly. Clearly, rapid claw closure must have evolved before the ability to snap, but its evolutionary origins are unknown. Unearthing the functional mechanics of transitional stages is therefore essential to understand how such radical novel abilities arise. We reconstructed the evolutionary history of shrimp claw form and function by sampling 114 species from 19 families, including two unrelated families within which snapping evolved independently (Alpheidae and Palaemonidae). Our comparative analyses, using micro-computed tomography (microCT) and confocal imaging, high-speed video, and kinematic experiments with select 3D-printed scale models, revealed a previously unrecognized “slip joint” in non-snapping shrimp claws. This slip joint facilitated the parallel evolution of a novel energy-storage and cocking mechanism—a torque-reversal joint—an apparent precondition for snapping. Remarkably, these key functional transitions between ancestral (simple pinching) and derived (snapping) claws were achieved by minute differences in joint structure. Therefore, subtle changes in form appear to have facilitated wholly novel functional change in a saltational manner.
Keywords: Alpheidae, Palaemonidae, innovation, functional morphology, biomechanics, evolutionary morphology, evo-devo, comparative morphology, saltational evolution, torque-reversal joint
 |
| Figure 1. MicroCT Images, Torque Moment Arms, and Schematic Illustrations of Three Shrimp Claw-Joint Types When Closed and Fully Open. (A) Pivot joint: anterior face∗ of right P1 in a basally branching caridean shrimp. (B) Simple slip joint (no torque reversal or power amplification): anterior face∗ of right P2 in an “intermediate” caridean shrimp. (C) Cocking slip joint (type 1 torque-reversal cocking, most likely power-amplified closing): anterior face∗ of right P1 in a feebly snapping alpheid shrimp. (A’–C’) Overlaid sagittal plane and surface rendering (via micro-computed tomography [microCT]) of claws of all three species showing torque moment-arms (+, –) when closed (upper) and fully opened (lower and background); negative torque (–) indicates that initial contraction of part of the closer muscle causes cocking. (A”–C”) Schematic representation of all three joint types showing loading orientations of opener and closer muscles. (A”) Pivot joint: purely rotational motion of dactyl. (B”) Slip joint: during opening, the dactylar base both rotates and translates (slips) across the propodal ridge (B). (C”) Cocking slip joint: during opening, the dactylar base both rotates and translates—including an abrupt sliding motion into the fully cocked position, where part of the closer muscle (gold) will generate reversed torque (–), and hence energy storage, because it inserts above the fulcrum (white dot). White dots show primary rotation axes (A–A”) or fulcrum points (B–B” and C–C”) for dactylar sliding and rotation. Black dots identify a reference point on the dactylar base. White arrows (A–C and A”–C”) show dactylar base trajectories during opening; closing would follow the same trajectories but in reverse. Red arrows (A’–C’) indicate dorsal-most closer-muscle contraction vectors (labeled V1 in Figure 4). Yellow arrows (A’–C’) represent torque moment arms about the fulcrum. Scale bars, 500 μm (A) and 300 μm (B and C). om, opener muscle; cm, closer muscle; (+), positive (counterclockwise) initial torque during claw closing; (–), negative (clockwise) torque during claw cocking generated by the gold-shaded muscle region in (C”). See also Figure SM1 in Methods S1 (joint-type scoring), Figures S1–S4 (microCT images of all claws), Movies S1A and S1B (actual dactyl motion), Movies S2A–S2E (3D model tests), and Table S1 (joint types of all species). ∗See Supplemental Results (Methods S1) for an explanation of claw-face viewing perspectives. |
Tomonari Kaji, Arthur Anker, Christian S. Wirkner and A. Richard Palmer. 2018. Parallel Saltational Evolution of Ultrafast Movements in Snapping Shrimp Claws. Current Biology. 28(1); 106-113. DOI: 10.1016/j.cub.2017.11.044
An adaptation 150 million years in the making phy.so/434192683 via @physorg_com